

Which Statement About Animal Behavior Is Most Accurate?
People take pondered beast behavior since the beginnings of man existence (Frison 1998). Insights into animal behavior immune our ancestors to outsmart prey during a hunt or befriend another brute, the latter eventually leading to domestication of animals. Knowledge of animal mating systems helped human societies grow, because propagation of domesticated stocks ensured that at that place were dependable supplies of food (Zeder 2008).
When observing wild or domesticated animals today, nosotros are ofttimes fascinated by their persistence in accomplishing a given task, such as when magpies adapt private sticks to form a nest or when a pet seeks out its favorite toy (Figure ane). And, we continue to ask basic questions near animal behavior: Exercise animals accept motivations and preferences? Do they make conscious choices? When given alternative partners or various foods to cull amidst, do animals make adaptive choices that are good for them? These and similar questions are addressed in controlled experimental designs that measure and test connections betwixt an observed behavior and its hypothesized affectors.

Figure 1
Pet dogs ordinarily enjoy play and can develop strong preferences for specific activities and toys.
What are Motivations, Preferences and Choices, and Why Practice Animals Accept Them?
Motivation, the desire or willingness to engage in a behavior, can be positive or negative and vary in strength. Motivation is positive for tasks and outcomes that animals wish for and negative for undesirable activities. Positive and negative motivations are inferred from behavior, for instance, from approach-behavior vs. avoidance, respectively (Kirkden & Pajor 2006).
Motivational strength varies between individuals. Some components of this variation can exist ascribed to constant individual factors like species-specific characteristics or gender, while others are explained by changing individual or ecology factors like age, feel, time of twenty-four hour period, conditions, and resource predictability. In other words, a motivation is generally influenced by many factors that tin be intrinsic (e.g., genetic or physiological) or extrinsic (i.due east., in the fauna's environment). To illustrate, the motivation to drink (thirst) can be increased past hormones responsible for controlling the trunk's h2o balance, only is also enhanced by the sight of water.
Preferences are based on an ability to evaluate sets of simultaneously bachelor alternatives that satisfy the same motivation and to aspire toward the one opportunity that is almost desirable. A preference may exist specific to the individual (such equally preferring potato chips over basics), and refers to the difference in motivational force to get the ane resources over the other, or others. Ultimately, animal preferences are inferred from choice behavior. Choice behavior refers to what an brute actually does — the consequences of its preferences and its final decisions.
In summary, earlier animals make choices they go through a decision-making process guided by their motivations and preferences. Presumably, this strategy is adaptive, insofar as being advantageous to the animal and favored by evolution via natural option: Individuals that choose benign options, similar eating the more nutritious food or resting at the safer nesting site, are also more than likely to have offspring that are successful in reproduction and survival (Existent 1991).
How Can We Assess What Animals Prefer and the Strength of Their Preferences?
It is feasible to measure beast choice behavior in the wild and make inference well-nigh what animals prefer. Jane Goodall, for example, used ad libitum behavior sampling to reveal details about the preferred placement of chimpanzees' sleeping-nests in Tanzania. She found that nests typically were built in rather inaccessible places, presumably as a predator abstention strategy (Goodall 1962). Thus, behavioral data from field studies can provide useful information about fauna choice behavior, but the results can as well be challenging to interpret considering many factors that influence behavior are outside the researcher's control.
Many scientists examine animal behavior in standardized test situations in the laboratory. In this setting, behavioral affectors are more easily monitored and controlled for than in the wild. For reliable command over experimental variables and optimal power to test behavioral affectors, laboratory studies are unremarkably performed with captive-born or domesticated animals that are familiar with semi-natural or artificial housing situations. In the laboratory, an fauna'southward preferences are tested by allowing it to cull between two or more defined options. Next, when the animal's choice beliefs reveals the pick that is preferred or avoided in the experiment, the scientist can move on to ask how strong its behavioral preference is. Such tests are termed motivation tests.
In laboratory testing of factors that can influence beliefs, treatment animals, which are exposed to the factor or factors of involvement, are assorted to reference observations. In some experiments, the reference observations are fabricated initially to make up one's mind the baseline behavior of each animal. After handling, the animal is compared to its own baseline as a reference or internal control. In other experimental designs, separate subsets of individuals form a reference or control group that parallels the treatment in all aspects other than the factor(s) of involvement. Subsequent comparisons of the control and treatments groups allow scientists to conclude about the influences on behavior, with fewer disturbances from intrinsic and extrinsic elements that can confound interpretation.
Instance Report i: Social Preference and Motivational Forcefulness
Amongst dogs, it is important for young animals to collaborate with others for play and social learning, and to feel safe. The same is true for other canids including captive silver foxes, known in the wild as the blood-red fox (Vulpes vulpes). At 2 months of historic period, fob pups prefer social contact and respond towards both familiar and unfamiliar pups in a playful manner. Notwithstanding, as the foxes age from pups to seven-calendar month-onetime juveniles, they spend less time with a social companion and their behavior towards strangers changes from playful to aggressive (Akre et al. 2009).
How can we measure whether juvenile foxes are motivated for social contact?
A method for testing an animal's strength of motivation is to ask it to "pay" an entrance fee for access to a resources and then measure the maximum "price" it is willing to pay. But what sort of currency can be used with animals like foxes? The currency is typically a learned behavior, similar pushing a heavy door or pulling a loop string, a so-called operant response. Besides, when using operant methodology to test whether social contact is of import; the cost foxes pay for access to a companion must be compared with access to a baseline resource of known importance to them. One such resource is food, since animals are motivated to satisfy their hunger (Jackson et al. 1999). The foxes' valuation of food thereby becomes the internal reference-behavior of this study.
This system (Effigy 2) was used to test the strength of social motivations in six juvenile female silver foxes that had been trained to pull a loop to open a door. When a fox delivered a required number of pulls, the door opened into a chamber. The foxes' motivations were examined in two split trials: During 1 trial, the foxes worked for food placed within the sleeping accommodation to found their individual reference value. During the other trial, the foxes could socialize with some other female when they visited the chamber. The companion could freely enter or leave, which insured that beingness together was voluntary.
In each trial, the experimenters raised the entrance fee daily by requiring more pulls earlier the door would open, until each creature had paid its maximum cost and ended its visit to the chamber.
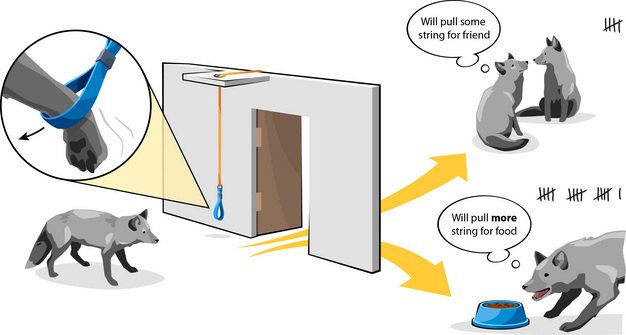
Effigy 2
An operant technique using the "maximum price approach" measured the strength of motivation for social contact vs. food in silver foxes. Each fob delivered increasing numbers of pulls on a loop string for access to food, the baseline resources. Numbers were compared to the corporeality of pulling that the same animals delivered for admission to a companion.
The researchers could now work out the average price the juvenile foxes were willing to pay for each resource. Nutrient was the near desired commodity with an average of 1632 pulls betwixt the six females (min 224, max 2368). However, the price paid for social contact averaged as much as 512 pulls (min 128, max 992), which represented 38% of the value that the foxes placed on food when hungry. Thus, the researchers concluded that female silvery foxes do desire social contact equally juveniles (Hovland et al. 2008).
Such operant experiments, relying on unproblematic behavioral responses that allow animals to show us what they desire, can give valuable insight into resources that are important for brute well-being.
Case Study two: Food Preference and the Physiology of Choice Behavior
Among beloved bees (Apis mellifera), individuals participate in social food-gathering exterior the colony. Some bees prefer protein-containing pollen from flowering plants, while others seek out nectar — a rich source of carbohydrates. Bees with different foraging preferences also bear witness different expression-levels of insulin-sensing genes in their abdominal fat (Amdam & Page 2010). Insulin sensing is active in muscles, liver and fat during uptake of sugar (glucose) from the claret, but insulin signals tin also human action remotely on the encephalon to regulate eating behavior (Vettor et al. 2002).
How can we test whether insulin-sensing genes in the bees' abdominal fat tin human action remotely on the brain, to affect behavioral preferences toward pollen and nectar collection?
Genetic effects on behavior tin be studied in laboratory animals where the genes of interest are "knocked downward", then their expression is disrupted. Similar factor knockdown tin exist achieved in the abdominal fat cells of honey bees with a technique called RNA interference. Using RNA interference, scientists compared 150 knockdown dearest bees toward a reference group with a similar number of control bees. The controls went through a matching technical and biochemical handling-procedure that did non crusade cistron knockdown. The knockdown treatment targeted IRS (the Insulin Receptor Substrate); a gene that is central to insulin sensing. The knockdown and control bees were tagged, transferred to beloved bee colonies, and advisedly observed. Tagged bees were captured when returning to their host colonies from foraging flights, and the researchers measured their gustatory (taste) response to saccharide besides as how much pollen and nectar the bees nerveless (for more details, see Wang et al. 2010).
The scientists found that the treatment and control bees responded equally to carbohydrate, but the IRS knockdown bees collected about xxx% less nectar and proportionally more pollen than the controls. The study concluded that an insulin-sensing cistron in the intestinal fat of love bees could influence the insects' foraging pick-beliefs without altering the bees' taste for saccharide (Wang et al. 2010).
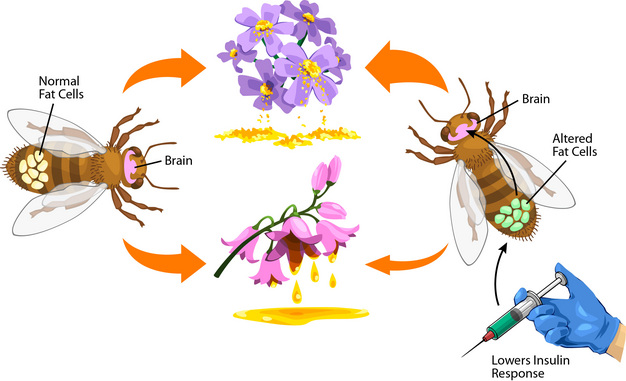
Figure 3
By reducing insulin sensing in abdominal fat cells of honey bees, scientists revealed a role of these cells in nutrient-choice beliefs. Afterward a key factor in insulin sensing was knocked down (Contradistinct Fatty Cells), the bees abandoned sugar-containing nectar in favor of protein-rich pollen. The control group (left) received equivalent technical and biochemical handling just retained normal fatty cells.
Such basic experiments on how physiology can bias individuals toward different food cravings tin can aid our progress in agreement food-related beliefs and how information technology influences human wellness.
Glossary
Adaptive behavior – Any behavior that enables an animal to accommodate to a specific situation or environment, and that supports the animal's long-term survival and reproduction.
Ad libitum beliefs sampling – To record all instances of behavior that seem advisable in an animal or in groups of animals. Requires no specific rules for how the behavior is recorded or which animals are recorded.
Controlling – To evaluate the perceived stimulus and select an appropriate behavioral response. The process of controlling can be simple (e.thou., rely on sensory filters), or involve circuitous cognitive processes like problem-solving.
Experimental controls – Subjects that practice not receive experimental handling only are similar to treated subjects in all other respects.
Experimental paradigm – Established methods that allow experiments to be conducted by specific standards. Typically rely on theory that guides methods-development and use.
Operant response – A learned behavior established based on the consequences of its operation.
RNA interference – Enzymatic cleavage of an mRNA based on its recognition by the RNA-induced silencing complex (RISC). Recognition uses a brusk template RNA in RISC that is gratis to a segment of the target mRNA sequence. RISC-mediate cleavage of mRNA can silence gene expression by reducing the amount of mRNA available for protein synthesis.
References and Recommended Reading
Akre, A. Grand., Bakken, Chiliad., & Hovland, A. Fifty. 2009. Social preferences in farmed silvery fox females (Vulpes vulpes): Does it change with historic period? Practical Brute Behaviour Science 120, 186–191 (2009).
Amdam, G. V. & Page, R. E. The developmental genetics and physiology of honeybee societies. Animal Behaviour 79, 973–980 (2010).
Frison, G. C. Paleoindian large mammal hunters on the plains of North America. Proceedings of the National Academy of Sciences of the Us of America 95, 14576–14583 (1998).
Goodall, J. Nest building behavior in complimentary ranging chimpanzee. Register of the New York Academy of Sciences 102, 455–467 (1962).
Hovland, A. Fifty. et al. The nature and force of social motivations in immature farmed silver play a trick on vixens. Applied Animal Behaviour Scientific discipline 111, 357–372 (2008).
Jackson, R. E., Waran, Northward. One thousand., & Cockram, One thousand. Southward. Methods for measuring feeding motivation in sheep. Animal Welfare 8, 53–63 (1999).
Kirkden, R. D. & Pajor, E. A. Using preference, motivation and disfavor test to ask scientific questions nearly animals' feelings. Applied Animate being Behaviour Science 100, 29–47 (2006).
Real, Fifty. A. Animate being choice beliefs and the evolution of cognitive compages. Science 253, 980–986 (1991).
Vettor, R. et al. Neuroendocrine regulation of eating behavior. Periodical of Endocrinological Investigation 25, 836–854 (2002).
Wang, Y. et al. Down-regulation of honey bee IRS gene biases beliefs toward food rich in poly peptide. PLoS Genetics six:e1000896 (2010).
Zeder M. A. Domestication and early agriculture in the Mediterranean Bowl: Origins, diffusion, and impact. Proceedings of the National Academy of Sciences of the United states 105, 11597–11604 (2008).
Source: https://www.nature.com/scitable/knowledge/library/measuring-animal-preferences-and-choice-behavior-23590718/
Posted by: pridgenforome.blogspot.com
0 Response to "Which Statement About Animal Behavior Is Most Accurate?"
Post a Comment